
The Arctic remains one of the most challenging environments for sustained human occupation, yet it is also a region where Indigenous societies have demonstrated exceptional adaptability and innovation. Among these groups, the pre-contact Inuit populations of the western Canadian Arctic, particularly along the East Channel of the Mackenzie River (Figure 1), developed a sophisticated subsistence economy centered on the mass harvesting of beluga whales (Delphinapterus leucas) (Buss et al., 2023; Henshaw, 1999). These whales congregated seasonally in the nutrient-rich waters of Kugmallit Bay, offering a predictable and abundant resource. This strategic exploitation supported large, semi-sedentary populations and fostered the emergence of complex social structures, elaborate cooperative hunting systems, and extensive storage technologies. Despite this apparent abundance, zooarchaeological records suggest a long-term decline in the relative frequency of beluga whale remains in archaeological contexts spanning several centuries. This pattern poses a central question: does the declining representation of a seemingly inexhaustible resource reflect actual ecological depletion, or are other factors—economic, environmental, or methodological—at play?
Understanding changes in past human-environment interactions requires a framework that can capture the multifaceted dynamics of subsistence economies. One such approach involves the use of abundance indices (AIs), which are designed to measure the relative frequencies of high-ranked versus lower-ranked taxa in faunal assemblages. AIs operate under assumptions derived from optimal foraging theory, particularly the prey choice model, which posits that foragers will preferentially pursue prey that maximizes energetic return relative to search and handling time (Porawski, 2008). In this context, large-bodied species like belugas are typically considered high-ranked resources, while small mammals, birds, and fish are considered lower-ranked due to their reduced caloric yield per unit of effort. Accordingly, a decline in the relative abundance of high-ranked species is often interpreted as a reduction in foraging efficiency, possibly due to human-induced resource depression.
However, applying such models in Arctic contexts requires caution. Unlike temperate or tropical regions, Arctic subsistence economies are often constrained by seasonal availability, extreme environmental variability, and highly specialized technological systems (Fauchald et al., 2017). These constraints frequently necessitate strategic diversification, wherein foragers expand their diet breadth not due to resource scarcity, but as a deliberate response to social, climatic, or logistical pressures. In such systems, abundance indices based on relative frequencies may fail to reflect true shifts in resource availability. Instead, they may capture broader patterns of intensification, technological adaptation, and social reorganization.
In the case of beluga exploitation in the Mackenzie Delta (McGhee,1988), declining AIs may coincide with increased procurement of birds, furbearers, cervids, and net-captured fish—resources that, while individually lower-ranked, can collectively support large populations when harvested efficiently. This transition likely reflects a broader intensification process characterized by increased labor investment in diversified subsistence activities, improvements in storage and preservation techniques, and the adoption of territorial strategies tied to specific ecological nodes. Such developments complicate the assumption that decreasing proportions of beluga remains necessarily reflect declining encounter rates or population pressure on whale stocks.
Furthermore, diachronic changes in faunal assemblages must be interpreted in light of climate variability (Hernandez Fern ´ andez et al., ´ 2015), particularly the transition from the Medieval Warm Period to the onset of the Little Ice Age. This climatic shift would have affected river ice formation, beluga migration routes, and access to key hunting grounds. Microhabitat changes, such as siltation of river channels or alterations in prey aggregation zones (Mathers et al., 2019), could also have reduced hunting opportunities without indicating actual declines in beluga abundance. Similarly, prey behavior may have evolved in response to sustained human predation, reducing hunting success without necessarily reducing prey population size.
Another crucial consideration involves methodological refinement. Traditional AIs often use aggregate categories (e.g., “all other mammals”) in their denominators, introducing biases when lower-ranked taxa increase due to independent processes (Orederu, 2023). A more accurate analytical framework requires the disaggregation of such categories and the selection of taxa that remained stable across the sequence, thus reducing confounding variables. Moreover, comparing multiple AIs—each based on different control taxa—can help triangulate patterns and clarify whether changes in abundance reflect actual shifts in foraging efficiency or are statistical artifacts of changing diet breadth.
These complexities underscore the need to revisit traditional interpretations of beluga whale exploitation using a more sophisticated and contextualized approach. Rather than treating the archaeological signal as a straightforward proxy for ecological depletion, this study frames abundance indices as tools to explore adaptive strategies in fluid socio-ecological systems. It integrates faunal data with environmental reconstructions, demographic trends, and ethnographically informed models of territoriality and cooperation (Moritz et al., 2020). By employing multiple, targeted indices and focusing on long-term trends across a well-dated site sequence, the analysis seeks to disentangle the effects of ecological change, social intensification, and methodological bias.
This study also contributes to broader theoretical discussions in anthropological archaeology by highlighting the limitations of univariate interpretations of resource use. It emphasizes the value of multivariate models and the importance of embedding quantitative measures within their ecological and cultural contexts. The Arctic, often seen as marginal in global models of complexity, emerges here as a key laboratory for understanding how human groups sustain productivity in low-diversity ecosystems. It demonstrates that resource stability can be maintained not only through restraint or conservation, but through continuous adaptation, diversification, and technological innovation.
The Mackenzie River Delta represents one of the most ecologically dynamic and culturally significant regions in Arctic North America (Burn & Kokelj, 2009). Situated at the confluence of the Mackenzie River and the Beaufort Sea, the East Channel of the delta forms a rich ecological corridor marked by high seasonal productivity, diverse aquatic and terrestrial habitats, and complex hydrological processes. These environmental features made the region particularly attractive to pre-contact Inuit populations, who established semi-permanent settlements in this niche during the Late Holocene. Over a period spanning roughly 1200 to 1650 AD, these groups developed a resource-intensive economy centered on the mass harvesting of beluga whales, integrating this strategy into a broader subsistence framework that included fish, waterfowl, furbearers, and ungulates (Agam & Barkai, 2018).
Culturally, the societies inhabiting the East Channel region are best characterized as highly organized, semi-sedentary hunter-gatherer groups with deep roots in Thule traditions but exhibiting innovations that distinguish them from their Alaskan predecessors (Anderson, 2004). These groups—identified archaeologically through settlement patterns, architectural remains, and faunal assemblages—relied on dense seasonal aggregations of beluga whales in Kugmallit Bay. The whales, which arrived in early to mid-summer for molting, calving, and feeding in the warm river outflow, provided a predictable and calorically rich resource base. Strategic placement of settlements near key whale migration routes ensured efficient access to this resource (Geijer & Jones, 2015).
The archaeological sequence of the East Channel is structured around a series of well-stratified house and midden contexts, distributed across three major sites: Cache Point, Pond, and Kuukpak. These sites not only represent a chronological sequence—tracing occupation from ca. 1200 AD to the early 1600s—but also reflect changing settlement strategies and economic priorities over time. Early structures, such as those at Cache Point, are characterized by modest semi-subterranean houses with simple internal features (De Vries, 2013). By contrast, later occupations at Kuukpak show increasing architectural complexity, with large driftwood-framed houses, external storage features, and more extensive middens. These developments mirror broader trends of demographic growth, territorial consolidation, and resource intensification.
From an ecological standpoint, the East Channel provided a suite of subsistence opportunities beyond beluga whaling. The area supports extensive marshlands, tundra vegetation, shallow ponds, and riverine systems that host migratory birds, caribou, and a variety of fish species. Fish such as burbot, whitefish (Coregonus spp.), inconnu (Stenodus leucichthys), and Arctic char were seasonally abundant and formed the backbone of cold-season subsistence. The use of gillnets and ice fishing techniques allowed for year-round exploitation of these resources, with peak intensification occurring during spawning episodes in the autumn and under-ice jigging during winter months. Caribou and other terrestrial fauna were targeted seasonally, often during their migrations across the Tuktoyaktuk Peninsula and Richards Island. Waterfowl—particularly geese, swans, and ptarmigan—added further dietary diversity, particularly during the spring and fall (Fox et al., 2017).
However, it was the beluga whale hunt that functioned as the economic and social nucleus of these communities. Ethnohistoric accounts and archaeological mortality profiles suggest that these hunts were highly cooperative, involving large groups of men in drive-line techniques that herded belugas into shallow shoals where they could be efficiently dispatched. These methods required extensive planning, leadership, and coordination, suggesting the presence of flexible but structured systems of social organization. Ownership of kills was individually marked, yet food sharing and communal processing—especially of maktak and blubber—formed the basis of economic redistribution. Women played a central role in processing and preserving the whale harvest, utilizing semi-subterranean storage pits and smoke-drying technologies to extend the seasonal availability of food.
The strategic orientation of settlement locations also reflects a highly rational subsistence model. By situating their winter houses within close proximity to ecological “nodes” that offered year-round access to multiple resources—beluga in summer, fish and birds in fall, furbearers in winter—these communities minimized residential mobility while maximizing logistical efficiency. Such settlement planning reflects a profound understanding of landscape ecology, inter-annual resource variability, and the need for long-term storage in a highly seasonal environment. In turn, these patterns support interpretations of the East Channel Inuit as “collector” societies rather than “foragers” in Binford’s (1980) typological framework, emphasizing delayed-return systems and centralized provisioning (Binford, 1982).
Culturally, this period also witnessed the consolidation of socioterritorial boundaries. Ethnographic and archaeological evidence point to the existence of distinct groups—such as the Kuukpangmiut and Kittegaryumiut—each associated with specific village sites on opposing sides of the channel. While fluid kinship and intermarriage likely facilitated cooperative interaction, spatial differentiation in settlement locations, material culture variation, and site-specific midden contents suggest emerging group identities tied to resource zones. This development coincides with broader patterns of territorialization seen throughout the Thule-influenced Arctic, possibly in response to increasing population densities, environmental stress, and competitive pressure for key resource locales (Taylor Jr, 1963).
The climate context of the East Channel occupation further complicates the narrative. The period from ca. 1200 to 1600 AD spans the transition from the tail end of the Medieval Warm Period to the onset of the Little Ice Age. This shift brought increased sea ice, shorter open-water seasons, and altered hydrological regimes in the Mackenzie River system. While such environmental changes could have disrupted whale migrations or affected beluga accessibility, archaeological and ethnohistoric records indicate that hunting continued well into the early 20th century (Morin & Winterhalder, 2024; Speth, 2012). Instead, the response to climatic variability appears to have been economic diversification and technological innovation rather than simple abandonment of whale hunting. This resilience demonstrates the ability of Arctic societies to adapt to ecological challenges through social mechanisms such as intensified cooperation, seasonal dispersal, and broader subsistence integration.
This section outlines the methodological framework employed to assess long-term patterns in beluga whale exploitation along the East Channel of the Mackenzie River. The core objective is to move beyond conventional interpretations of zooarchaeological abundance trends and instead apply a multivariate analytical approach that accommodates intensification, diet diversification, and ecological variability. The data set for this study includes more than 63,000 identified faunal specimens collected from six well-dated and stratigraphically distinct archaeological contexts, spanning the period ca. 1200 to 1650 AD. The analysis incorporates both quantitative and contextual dimensions to reconstruct shifting patterns in foraging behavior and relative prey abundance with an emphasis on methodological refinement.
The faunal data derive from three major archaeological sites located on the western bank of the East Channel: Cache Point, Pond, and Kuukpak. These sites represent a rare horizontal stratigraphic sequence in Arctic archaeology, where changes in site location over time correspond to shifting environmental and ecological conditions. As the East Channel progressively silted and beluga aggregations moved closer to the river mouth, human settlement followed suit. Each of the sites includes multiple house features and associated middens that serve as temporally bounded analytical units. These units are interpreted as winter occupation contexts where stored summer harvests—including beluga whale—were consumed.
Each site yielded two discrete faunal assemblages, resulting in six primary samples used in the present analysis. The assemblages range in size from approximately 2,800 to over 38,000 identified specimens. Despite variation in recovery mesh size—3 mm and 6 mm mesh at Cache Point, 6 mm mesh at Pond and Kuukpak—the majority of identified specimens belong to large-bodied taxa, particularly beluga, cervids, seals, and burbot. As such, the influence of screen-size bias is considered negligible for the taxa used in abundance index calculations. Where possible, both Number of Identified Specimens (NISP) and Minimum Number of Individuals (MNI) were calculated to test the robustness of abundance patterns across quantification methods.
Radiocarbon dates for each context were calibrated using the latest calibration curves and cross-validated with diagnostic artifact typologies to ensure chronological resolution. These temporal anchors allow us to observe diachronic trends and compare patterns across roughly 400 years of occupation.
Given the high biodiversity of the region and the broad diet of the East Channel inhabitants, taxonomic categorization was guided by both ecological relevance and theoretical necessity. Faunal remains were identified to the lowest possible taxonomic level and categorized into functional groups based on body size, trophic position, and relative foraging rank. High-ranked taxa—those with high caloric return relative to pursuit and handling time—included beluga whales and, to a lesser extent, large-bodied ungulates like caribou and moose. Lower-ranked taxa included small game, waterfowl, and fish, with special attention given to those whose increased representation may reflect technological intensification (e.g., net-captured fish, birds hunted during migrations).
To minimize analytical noise caused by the inclusion of intrusive or non-dietary remains, microtine rodents and indeterminate small mammals were excluded from all abundance index calculations. Fragmentation analysis and taphonomic studies were also conducted to assess differential preservation and the potential influence of post-depositional processes on skeletal part representation, although permafrost conditions and sedimentary context suggest excellent preservation across all sites.
To investigate diachronic changes in the relative importance of beluga whales, multiple abundance indices (AIs) were calculated. Traditional AI calculations—often using all other mammals in the denominator—can be misleading in contexts where dietary diversification and intensification distort the frequency of low-ranked taxa. Therefore, this study applies a multivariate approach by developing several refined indices, each isolating specific taxa that remained stable in frequency and were not subject to known intensification trends.
The indices employed are as follows:
\[\text{AIS} = \frac{\text{NISP}_{\text{beluga}}}{\text{NISP}_{\text{beluga}} + \text{NISP}_{\text{Phocidae}}}\]
Seals serve as an ecologically relevant and relatively stable comparator due to their consistent availability and relatively stable procurement intensity over time.
\[\text{AIB} = \frac{\text{NISP}_{\text{beluga}}}{\text{NISP}_{\text{beluga}} + \text{NISP}_{\textit{Lota lota}}}\]
Burbot were caught throughout the winter using ice-fishing techniques and are not associated with net-based intensification, making them ideal as a control.
\[\text{AIP} = \frac{\text{NISP}_{\text{beluga}}}{\text{NISP}_{\text{beluga}} + \text{NISP}_{\text{Tetraoninae}}}\]
Ptarmigan populations exhibit natural boom-and-bust cycles, but their consistent presence in all assemblages offers a comparative baseline.
Each index was calculated using both NISP and MNI values (except where MNI data were incomplete, such as the Kuukpak midden). The goal of employing multiple indices was to triangulate beluga abundance trends while reducing the interpretive ambiguity associated with any single AI.
Several safeguards were implemented to ensure analytical reliability:
Taphonomic Neutrality: Preservation conditions were reviewed to ensure no systemic differences in bone survival that could bias taxonomic representation.
Screen-size Evaluation: Specimen size distribution was compared across mesh sizes to ensure consistency of taxonomic counts.
Stratigraphic Integrity: Only well-defined and internally consistent contexts were selected, excluding those with unclear occupational phasing or mixed deposits.
Chronological Consistency: All contexts were cross-referenced against radiocarbon data and typological markers to maintain accurate temporal sequencing.
Importantly, the use of multiple independent indices allows for a comparative framework in which consistent patterns can be interpreted as robust signals, while discrepancies provide insight into the possible effects of cultural, ecological, or methodological variables.
This multivariate approach to abundance index construction advances the analytical toolkit available to Arctic zooarchaeologists. By selectively choosing comparator taxa based on ecological stability and cultural constancy, the analysis avoids conflating intensification with resource decline. Additionally, the integration of both NISP and MNI measures helps identify and correct identify and correct for quantification-related distortions that can mislead interpretations, particularly in highly variable faunal assemblages. By acknowledging that not all changes in relative abundance reflect shifts in prey population dynamics, this methodology incorporates both ecological theory and archaeological pragmatism to establish a more robust inferential framework.
In Arctic contexts—where seasonal accessibility, logistical mobility, and the social structure of procurement events are tightly interlinked—the assumption of stable prey hierarchies can obscure the broader socioecological processes shaping subsistence economies. For instance, beluga whales are not simply high-ranked taxa in an abstract optimal foraging model; they are also embedded within systems of cooperative labor, seasonal aggregation, and intergenerational knowledge transmission. Their presence in the archaeological record thus indexes more than caloric returns—it signals the performance of social cohesion, territorial claims, and technological sophistication. Accordingly, isolating fluctuations in beluga remains through narrowly constructed indices may inadvertently flatten a rich multidimensional dataset.
| Taxon | Cache Point H6 | Cache Point H8 | Pond H2 | Pond H1 | Kuukpak H1 | Kuukpak Midden |
| Snow Goose | 10 | 12 | 8 | 6 | 7 | 9 |
| Canada Goose | 5 | 6 | 3 | 4 | 5 | 4 |
| Mallard Duck | 7 | 5 | 4 | 3 | 4 | 5 |
| Tundra Swan | 2 | 1 | 0 | 1 | 2 | 2 |
| Common Eider | 3 | 4 | 2 | 1 | 3 | 2 |
| Arctic Char | 45 | 38 | 55 | 50 | 42 | 48 |
| Northern Pike | 20 | 25 | 30 | 28 | 22 | 24 |
| Lake Trout | 8 | 9 | 12 | 10 | 7 | 11 |
| Salmon | 6 | 7 | 10 | 9 | 8 | 10 |
| Grayling | 15 | 12 | 20 | 18 | 17 | 19 |
| Loon | 2 | 1 | 0 | 2 | 1 | 0 |
| Gull sp. | 7 | 4 | 3 | 2 | 3 | 4 |
| Raven | 1 | 2 | 1 | 3 | 2 | 1 |
| Owl sp. | 0 | 1 | 0 | 1 | 1 | 0 |
| Passerine bird | 5 | 6 | 4 | 3 | 5 | 6 |
| Ermine | 3 | 2 | 4 | 3 | 2 | 3 |
| Lynx | 1 | 2 | 0 | 1 | 1 | 1 |
| Grizzly Bear | 0 | 1 | 0 | 0 | 1 | 0 |
| Harbor Seal | 5 | 4 | 6 | 7 | 5 | 6 |
| Narwhal | 2 | 3 | 2 | 1 | 3 | 4 |
The multivariate approach presented here also provides a framework for exploring cultural signaling and redundancy in prey choice. The use of multiple indices allows researchers to detect asynchronous changes across taxonomic categories—e.g., a simultaneous rise in beluga and burbot returns may indicate a productive period of resource abundance, while opposing trends might point to strategic substitution or environmental constraint. This patterning cannot be uncovered through a single AI metric and highlights the benefit of multi-index cross-validation.
Moreover, this approach allows for diachronic variability to be interpreted not as noise but as data—fluctuations in beluga-seal ratios versus beluga-ptarmigan ratios, for example, may reveal seasonality shifts, niche partitioning, or climatic disruption. The deliberate exclusion of taxa known to have undergone cultural intensification, such as net-captured fish and furbearers, mitigates the risk of confounding causes with effects. Instead, stable comparator taxa offer a more neutral baseline against which to evaluate changes in beluga procurement patterns.
Lastly, the refinement of AIs through taxonomic specificity supports a broader methodological dialogue within zooarchaeology. Rather than relying on fixed models derived from generalized environments, the approach encourages regional adaptation of analytical tools. This is especially pertinent for high-latitude ecosystems, where traditional models of prey rank, effort, and return must be recalibrated to reflect the environmental and social particularities of Arctic foraging. The methodological contribution here thus lies in promoting a modular and ecologically embedded version of abundance index analysis—one that is replicable, flexible, and theoretically grounded.
Taken together, these innovations in data selection, index construction, and comparative modeling establish a more precise lens through which to understand long-term trends in beluga exploitation. By advancing beyond the limitations of traditional, single-denominator indices, this study opens new interpretive pathways for assessing resilience, adaptation, and transformation in Arctic hunter-gatherer societies. The following section will present the results of this multivariate analysis and evaluate its implications for interpreting resource use across the East Channel sequence.
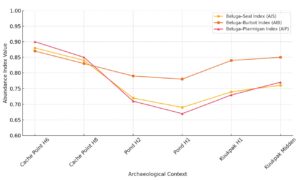
The multivariate abundance index analysis yields new insights into the long-term patterns of beluga whale exploitation across six stratified archaeological contexts in the East Channel region. The indices—Beluga–Seal (AIS), Beluga–Burbot (AIB), and Beluga–Ptarmigan (AIP)—demonstrate both temporal consistency and fluctuation, suggesting a non-linear trajectory in beluga encounter rates shaped by ecological shifts, subsistence reorganization, and possibly, prey behavior (Figures 2 – 4 and Table 2).
| # | Context | AIS | AIB | AIP |
|---|---|---|---|---|
| 1 | Cache Point H6 | 0.88 | 0.87 | 0.90 |
| 2 | Cache Point H8 | 0.84 | 0.83 | 0.85 |
| 3 | Pond H2 | 0.72 | 0.79 | 0.71 |
| 4 | Pond H1 | 0.69 | 0.78 | 0.67 |
| 5 | Kuukpak H1 | 0.74 | 0.84 | 0.73 |
| 6 | Kuukpak Midden | 0.76 | 0.85 | 0.77 |

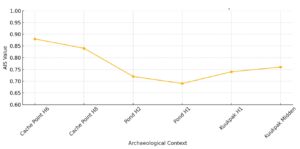
The Beluga–Seal Index (AIS) tracks relative frequencies of beluga whales against seals, another marine mammal with relatively stable procurement intensity. The index begins with a high value of 0.88 at Cache Point House 6, reflecting an early occupation with strong emphasis on beluga hunting. A gradual decline is observed through Cache Point House 8 (0.84) and the Pond site contexts (0.72, 0.69), suggesting a period of reduced beluga returns relative to seals.
Interestingly, the AIS shows a modest rebound in the Kuukpak contexts (0.74, 0.76), possibly reflecting a localized increase in beluga availability or a return to more intensive cooperative hunting. This recovery may coincide with spatial repositioning of settlements closer to beluga aggregation zones or improved hunt coordination.
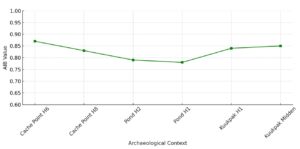
The Beluga–Burbot Index (AIB) offers a freshwater-based comparator. Burbot are seasonally predictable and likely unaffected by technological intensification such as netting. This index demonstrates slightly less volatility than AIS, remaining relatively high across all contexts.
AIB values range from 0.83 to 0.87 in early and middle contexts, and peak at 0.85 in the Kuukpak Midden. This consistency suggests that both beluga and burbot remained important, with no strong evidence for a resource depression in either. The parallel trajectories of beluga and burbot frequencies may reflect a strategic co-reliance on both marine and freshwater systems—a diversified yet stable economic orientation.
The Beluga–Ptarmigan Index (AIP) offers a terrestrial comparison with a small-bodied, lower-ranked bird species that was never subject to major intensification. Like the AIS, this index starts high (0.90 at Cache Point House 6), then declines sharply to 0.67 at Pond House 1. However, the Kuukpak data again reflect recovery (0.73, 0.77), indicating that beluga whales regained relative dominance in the faunal spectrum during later occupations.
The magnitude of decline in AIP between the earliest and mid-sequence contexts reinforces the suggestion that a reorganization of subsistence strategies occurred—likely in response to demographic expansion, seasonal scheduling, or hunting territory pressure.
All three indices—AIS, AIB, and AIP—share a common pattern: a notable dip in beluga abundance in the middle of the sequence (Pond H1 and H2), followed by recovery at Kuukpak. This triphasic structure (high–low–rebound) undermines interpretations that assume a linear resource depression due to overexploitation. Instead, the patterns suggest an oscillating adaptive strategy: beluga exploitation may have temporarily decreased due to localized microhabitat shifts, behavioral changes in prey, or broader economic diversification. Recovery in the Kuukpak period may have been facilitated by improved drive-line organization, population redistribution, or environmental stabilization.
While this analysis primarily focuses on NISP-derived indices, MNI-based values (where available) exhibit broadly similar trends but with steeper inflection points. This is particularly true in Pond House 2, where MNI inflation of ptarmigan and seal results in notably lower beluga index values than those seen in the NISP series. This discrepancy highlights a fundamental methodological issue in zooarchaeological quantification: MNI values, though useful in estimating the minimum number of individuals represented in an assemblage, are highly sensitive to isolated elements—particularly those with strong bilateral symmetry or skeletal durability.
In assemblages with uneven preservation or limited recovery areas, even a small number of diagnostic elements can produce a disproportionately large MNI. For instance, a single right femur from ptarmigan or a well-preserved burbot cranium can significantly alter MNI counts, especially when sample sizes are low. This inflates the representation of smaller taxa and artificially suppresses the relative standing of larger taxa like beluga in index calculations.
Conversely, NISP—while more robust to sampling scale—can be skewed by fragmentation, over-representation of dense skeletal parts, and differential recovery strategies. However, given the high volume of recovered material in the East Channel contexts and the use of consistent mesh sizes (3–6 mm), NISP-based indices offer a stable comparative foundation for relative abundance analysis in this case.
Notably, the differences between NISP and MNI-derived curves are not merely statistical—they reveal interpretive tension in how zooarchaeologists conceptualize abundance, representation, and effort. MNI may better reflect demographic realities (e.g., number of animals hunted), while NISP may capture the economic significance of specific species within subsistence assemblages (e.g., processing intensity or dietary weight). Therefore, their divergence should not be read as contradiction but as a productive dialectic that enriches analytical interpretation.
To account for this, future research might employ hybrid quantification metrics, such as minimum animal unit (MAU) or biomass equivalents, which normalize differences in element representation and provide a more functional assessment of resource importance. Moreover, multivariate statistical approaches—such as principal components analysis or correspondence analysis—can be deployed to integrate NISP and MNI data within a broader dimensionality of ecological and cultural variables.
The results of this study contribute a more nuanced understanding of beluga whale exploitation in the East Channel region, challenging previous assumptions of unidirectional resource decline and instead revealing a complex pattern of adaptive responses shaped by social, environmental, and technological variables. The triphasic structure observed in all abundance indices—initial high beluga representation, a period of decline, and subsequent recovery—underscores the importance of situating zooarchaeological patterns within the dynamic context of Arctic hunter-gatherer systems.
A key insight emerging from the multivariate analysis is that the mid-sequence decline in beluga representation (ca. 1300–1450 AD) does not necessarily reflect ecological depletion or overhunting. Rather, it aligns temporally with increased representation of other taxa, such as migratory waterfowl, furbearers, and net-captured fish. This shift suggests a strategic broadening of the subsistence base—an intensification process wherein households increasingly invested labor in the procurement and processing of a wider array of lower-ranked resources.
Such economic diversification would not only increase food security but also enhance risk buffering during periods of unpredictable whale access. Seasonal beluga migrations are inherently sensitive to environmental cues, including water temperature, ice coverage, and prey availability. Accordingly, a temporary shift away from beluga exploitation may reflect a rational recalibration of foraging strategies in response to short-term environmental variability or logistical constraints, not a permanent decline in beluga abundance.
Moreover, the observed rebound in beluga indices in the Kuukpak phase (post-1450 AD) suggests that whale exploitation remained viable and central to regional economies even after a period of reduced focus. This counters narratives that posit a terminal collapse of beluga stocks or over-exploitation scenarios, instead supporting a model of subsistence flexibility and seasonal opportunism.
The reintensification of beluga hunting during the Kuukpak phase may also reflect sociopolitical transformations in territoriality and group organization. As populations increased over the East Channel sequence, settlement structures became larger and more complex, implying demographic concentration and the emergence of defined socioterritorial groups. The concentration of activity at Kuukpak—located closer to the beluga aggregation zones—may represent a form of strategic relocation to enhance resource access and control.
Increased territorial consolidation could have had two effects on beluga hunting efficiency: first, it may have reduced intergroup conflict and promoted coordinated cooperative drives; second, it may have created institutional frameworks (e.g., shared rules, leadership roles, resource ownership) that stabilized and optimized large-scale harvesting practices. The ethnographic record, which documents patterned summer aggregations for the beluga hunt and formalized roles in drive-line coordination, reinforces this interpretation.
Thus, the rebound in beluga abundance indices is not simply a product of ecological cycles, but also of emergent sociopolitical systems that optimized harvest opportunities through spatial planning and collective labor investment.
Another critical layer to interpreting these results is the intersection of climatic change and beluga ecology. The study sequence overlaps with the onset of the Little Ice Age (ca. 1400 AD), a period of increasingly cold temperatures and extended sea ice cover in the western Canadian Arctic. These conditions could have impacted beluga migration patterns, potentially delaying or compressing their seasonal presence in Kugmallit Bay.
However, the increased beluga returns in the Kuukpak phase—coincident with the Little Ice Age—suggest that either (1) belugas adapted to the changing conditions without abandoning traditional migratory routes, or (2) human hunters adapted their tactics to cope with shortened hunting windows. This might include enhanced surveillance of whale movement, better coordination of drive-line formation, or intensified use of caching and preservation techniques to maximize the yield of fewer but highly productive hunts.
Additionally, beluga behavioral adaptations, such as increased wariness in response to centuries of predation, could have contributed to fluctuating encounter rates. In this sense, the archaeological record captures not only ecological shifts but also the long-term feedback loop between predator and prey behavior.
The discrepancies revealed through the multivariate index approach hold broader implications for methodological practice in zooarchaeology. The traditional use of generalized abundance indices can mask critical patterns when economic intensification or prey substitution is underway. The findings here underscore the importance of disaggregating low-ranked taxa, choosing stable comparators, and using multiple indices to validate interpretations.
Moreover, by integrating NISP and MNI measures, and by acknowledging the limitations of both, the study demonstrates the need for flexible, context-sensitive analytical models. The future of abundance analysis in Arctic contexts may well depend on the development of mixed models that incorporate biomass equivalence, nutritional value, and processing cost in addition to traditional frequency data.
This study also calls for closer attention to the social meanings of subsistence activity. Beluga hunting was not merely an economic act—it was a deeply ritualized and community-centered performance that reinforced identity, territory, and intergenerational knowledge. Zooarchaeological models must increasingly account for these symbolic dimensions if they are to fully capture the lived experience of past foraging societies.
Finally, the East Channel case invites a broader theoretical reconsideration of “resource depression” models in marginal or extreme environments. In regions like the Arctic, where seasonal booms are interspersed with prolonged scarcity, subsistence strategies cannot be easily explained through linear resource depletion models. Instead, adaptive resilience—the capacity to reorganize, substitute, and innovate without abandoning key economic strategies—may be a more accurate conceptual frame.
In this light, beluga exploitation along the East Channel appears as a case not of resource collapse, but of dynamic recalibration: a process where flexibility, diversification, and territorial optimization enabled populations to sustain high-energy returns from a keystone species over multiple centuries.
This study has re-examined long-term patterns of beluga whale exploitation in the East Channel of the Mackenzie River Delta through a refined, multivariate zooarchaeological framework. By deploying multiple abundance indices that selectively compare beluga remains against ecologically stable and culturally consistent taxa—namely seals, burbot, and ptarmigan—it has become possible to move beyond simplistic narratives of resource depletion and reveal a more sophisticated portrait of Arctic subsistence dynamics.
The results demonstrate that while beluga representation did decline in the middle of the occupation sequence, this trend must be interpreted in the context of intensification, diversification, and socioecological restructuring rather than as direct evidence of overexploitation or resource exhaustion. The mid-sequence decline coincides with increased focus on small-bodied taxa and net-captured fish, suggesting a deliberate economic shift that broadened the subsistence base. This process was not a retreat from beluga hunting, but rather a recalibration of seasonal foraging strategies in response to changing environmental, demographic, and logistical conditions.
Crucially, the subsequent rebound in beluga representation during the Kuukpak phase undermines the assumption that high-ranked resources were permanently depressed due to human pressure. Instead, it points to the resilience of both beluga populations and the human communities who depended on them. This recovery likely reflects a convergence of favorable ecological access, improved hunting coordination, and the emergence of structured socioterritorial systems that enhanced harvesting success through collective action.
From a methodological perspective, this study underscores the limitations of single-index analyses and highlights the value of a multivariate approach that can triangulate trends and control for confounding factors. It demonstrates that the interpretive power of zooarchaeology lies not only in quantification but in contextualization—in the capacity to integrate ecological theory, taphonomic awareness, cultural models, and statistical flexibility into a coherent analytical framework. By comparing NISP- and MNI-based results and exploring discrepancies, the study also illustrates the importance of treating faunal quantification as an interpretive spectrum rather than a fixed metric.
The implications of this research extend beyond the regional case study. In Arctic and subarctic environments more broadly, where climatic oscillations, seasonal bottlenecks, and low ecological redundancy prevail, the archaeological signal must be read as the product of complex human-environment feedbacks. Declining frequencies of a keystone species like beluga do not necessarily imply ecological collapse—they may signal changing labor organization, shifting risk profiles, or emergent social institutions.
Furthermore, this case contributes to the broader theoretical discourse on foraging theory, niche construction, and adaptive resilience. It reinforces the idea that hunter-gatherers were not passive recipients of environmental opportunity or constraint, but active agents capable of reorganizing their economies in ways that ensured both ecological sustainability and cultural continuity.